مکانیسم مقاومت داروئی در باکتریها
بدون شک مهمترین آنتیبیوتیکها، آنتیبیوتیکهای مهار کننده سنتز دیواره سلولی هستند. دسته مهم این آنتیبیوتیکها شامل آنتیبیوتیکهای بتالاکتام میباشد که شامل کارباپنمها، سفالوسپورینها، سفامایسینها، منوباکتامها و پنیسیلینها هستند. امروزه مقاومت به این آنتیبیوتیکها بسیار رایج بوده بطوریکه اغلب عوامل ایجاد کننده عفونتهای بیمارستانی به بیشتر این داروها مقاوم شدهاند. با توجه به این که کنترل و درمان عفونتهای ناشی از این باکتریها مستلزم آشنائی با مکانیسمهای ایجاد کننده مقاومت میباشد در این مقاله و مقالات بعدی اشارهای کوتاه به نحوه مقاومت باکتریها به آنتیبیوتیک میگردد.
مقاومت به آنتیبیوتیکهای بتالاکتام
مقاومت اکتسابی باکتریها به آنتیبیوتیکهای بتالاکتام احتمالاً از طریق جهش ژنهای کروموزومی یا انتقال از راه پلاسمید بدست آمده است. عموماً تغییرات وابسته به کروموزوم در حساسیت به آنتیبیوتیکهای بتالاکتام شامل تغییر در نفوذ پذیری آنتیبیوتیک یا تغییر در توانایی پروتئینهای باند کننده به پنیسیلینها (PBP) برای شناسایی آنتیبیوتیکها میباشد، اما بعضی از بتالاکتامازها توسط کروموزوم کد میشوند. مقاومت کد شده توسط پلاسمید نسبت به بتالاکتامها، همیشه بصورت معرفی یک ژن بتالاکتاماز میباشد.
- تغییر در نفوذ پذیری باکتری
نفوذ پذیری باکتریها نسبت به بتالاکتامها، ممـــــــــــــکن است در اثر تغییر در پورینها یا تغییر در لیپوپلیساکاریدهای غشاء خارجی، کاهش یابد. غشاء خارجی باکتریهای گرم منفی بصورت غیر قرینه است، یعنی پورینها و لیپوپلیساکارید در قسمت خارجی آن قرار دارند. اشریشیا کلی حدود۱۰۵× ۱ مولکول پورین در هر سلول دارد که میتواند بصورتهای کانالهای بزرگ (OmpF) یا کوچک (OmpC) باشد. در ابتدا، وقتی که غشاء خارجی اشریشیا کلی دارای کانالهای OmpC شده است، کاربنیسیلین و سایر داروهایی که معمولاً به آهستگی نفوذ میکنند، بطور اساسی قادر به ورود به باکتری نیستند. برخلاف سویههای اشریشیا کلی، سویههای انتروباکتر کلوآکه و سودوموناس ائروژینوزا به وضوح با تغییر در ترکیب لیپوپلی ساکاریدشان در مورد حساسیت به پنیسیلین با اشریشیا کلی تفاوت دارند.
(۲) تغییر در افینیتی اهداف آنتی بیوتیک
دومین مکانیسم مقاومت به آنتیبیوتیکهای بتالاکتام شامل تغییر در تمایل PBPها نسبت به آنتیبیوتیکهای بتالاکتام میباشد. بزرگترین ضربه کلینیکی حاصل از این مکانیسم، ظهور استافیلوکوک اورئوسهای مقاوم به پنیسیلین بوده است. سایر باکتریهایی که مقاومتشان نسبت به بتالاکتامها بدلیل تغییر در PBP است عبارتند از: کلستریدیوم پرفرنژنز (مقاومت به تمام پنیسیلینها)، هموفیلوس انفلوانزا و سراشیا (مقاومت به سفالوسپورینهای نسل سوم و بعضی از پنیسیلینها) و نایسریا گونوره آ و استرپتوکوک پنومونیه (مقاومت به پنی سیلین G).
- نقص در القاء اتولیز
بنظر میرسد که بعضی از باکتریها نسبت به بتالاکتامها تحمل پیدا کردهاند. آنتیبیوتیکهای بتالاکتام در این باکتریها نمیتوانند آنزیمهای اتولیتیک القاء کنند و نسبت MBC به MIC بیشتر یا مساوی ۳۲ میباشد. این بدان معنی است که پنیسیلین یا سفالوسپورین بیشتر از اینکه باکتریوسید باشد، باکتریواستاتیک است، مگر اینکه میزان بسیار زیادی آنتیبیوتیک مصرف شود. این امر در درمان افراد با ایمنی سرکوب شده و یا بیماران نوتروپنی مشکل ایجاد میکند. باکتریهایی که گاهگاهی تحمل به بتالاکتامها را نشان میدهند عبارتند از: لیستریا منوسیتوژنز، استافیلوکوک اورئوس، استافیلوکوک اپیدرمیدیس، استرپتوکوک آگالاکتیه و استرپتوکوک سانگوئیس؛ تمام این باکتریها گرم مثبت میباشند.
- تأثیر بتالاکتامازها
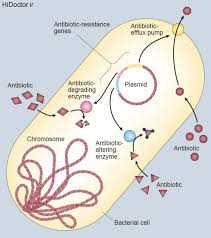
پردردسرترین مکانیسمهای مقاومت به آنتیبیوتیک، متعلق به بتالاکتامازها بوده است. یک بتالاکتاماز، آنزیمی است که بصورت غیرکووالانت به آنتیبیوتیکهای بتالاکتام باند میکند و با تشکیل یک پیوند کووالانت، سبب هیدرولیز باند آمیدی حلقوی حلقه لاکتام و آزاد شدن آنتیبیوتیک تغییر یافته (غیرفعال) میگردد. بتالاکتامازها برای آنتیبیوتیکهای بتالاکتام، با PBPها رقابت میکنند. امروزه بیشتر از ۱۰۰ نوع بتالاکتاماز شناخته شده که هر کدام الگو، اختصاصیت و تمایل مربوط به خود را دارند. اگر چه بتالاکتامازها در بین باکتریهای گرم مثبت و منفی بسیار پخش شدهاند، اما بنظر میرسد که توزیع آنها به صورت چند دسته بندی بزرگ است که دو نوع طبقه بندی زیر بسیار معمول میباشد.
اولین سیستم طبقه بندی مولکولی Ambler میباشد که براساس همولوژی پروتئین یا تشابه آمینواسیدی این پروتئینها، بتالاکتامازها را به ۴ کلاس اصلی دسته بندی میکنند که عبارتند از A، B، C و D (شکل ۱). در این نوع طبقه بندی ویژگیهای فنوتیپی مد نظر قرار نمیگیرد. در این نوع طبقه بندی، کلاسهای A، C، D بتالاکتامازهای حاوی سرین میباشند، در مقابل آنزیمهایی که در کلاس B قرار گرفتهاند، در واقع متالوبتالاکتامازها هستند که برای فعالیت خود نیاز به فلز روی دارند. این سیستم زیرمجموعه سیستمهای M.H.Richmond R.B.Sykes میباشد که در آن طبقه بندی بر اساس نوع سوبسترا، ژنهای کد کننده، بیان ژن و وزن مولکولی میباشد.
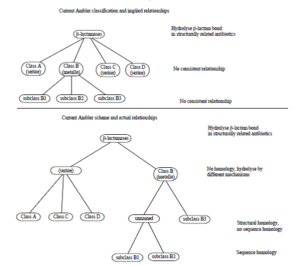
شکل ۱: دیاگرام شماتیک سیستم طبقه بندی آمبلر
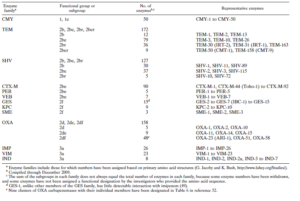
سیستم طبقه بندی رایج دیگر در خصوص بتالاکتاماز ها سیستم Bush jacoby medeiros میباشد که در آن بتالاکتامازها براساس نوع سوبسترا و مطالعات مهاری طبقه بندی میشوند (جدول ۱ و ۲). بتالاکتامازهای باکتریهای گرم مثبت که در این سیستم در یک کلاس واحد قرار دارند، آنزیمهایی هستند که به مقدار زیاد تولید شده و به محیط خارج ترشح میشوند، بطوریکه در خارج سلول میتوانند بتالاکتامها را غیرفعال کنند. دراین حالت، آنها از باکتریهایی که خودشان بتالاکتاماز تولید نمیکنند، نیز حفاظت میکنند. این پدیده اثر تلقیح (Inoculum effect) نامیده میشود و به هر آنزیم غیر فعال کننده آنتیبیوتیکی که توسط باکتریهای مقاوم به محیط ترشح میشود، مرتبط میباشد. در باکتریهای گرم منفی، بتالاکتامازها به شش کلاس اصلی تقسیم میشوند.
احتمالاً مهمترین بتالاکتامازهای سیستم ریچموند- سایکس در کلاس I قرار دارند. بتالاکتامازهای کلاس I توسط کلاوولانات یا سولباکتام مهار نمیشوند و بعضی از آنها، سفالوسپورینهای نسل سوم را میتوانند هیدرولیز نمایند. بیشتر بتالاکتامازها ساختمانیاند، اما بتالاکتامازهای استافیلوکوکی و کلاس I توسط سفامندول، سفوکسیتین و ایمیپنم القاء میشوند. بتالاکتامازهای کلاس II تا کلاس IV همگی توسط کلاولانات یا سولباکتام مهار میشوند. این بتالاکتامازها به میزان کم تولید شده و محدود به فضای پریپلاسمیک میباشند. بنابراین نفوذ پذیری غشاء خارجی بسیار بر عملکرد آنها اثر میگذارد و اثر تلقیحی نیز وجود ندارد. اگر تغییری در پورین روی دهد که سبب کم شدن نفوذپذیری شود، تأثیر بتالاکتامازها احتمالاً به میزان زیادی افزایش مییابد.
جدول ۱: انواع بتالاکتامازها در باکتریها
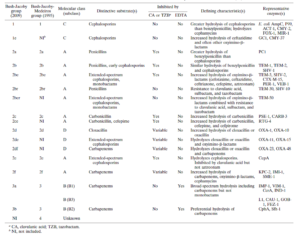
جدول ۲: خانوادههای مهم بتالاکتامازها از نظر بالینی