بیرجینگ دانشمند هلندی اولین بار کلمه ویروس (Virus) که به زبان لاتین به معنای سم است را بکار برد. در سالهای قبل از قرن نوزده، محققین مشاهده کردند عوامل بعضی از بیماریها از باکتریها کوچکترند که عوامل قابل پالایه شدن توصیف شدند. در سال ۱۸۹۲ برای اولین بار انتقال بیماری موزائیک تنباکو از سوسپانسیونی که از پالایه جداکننده باکتریها عبور داده شده بود، توسط ایوانووسکی نشان داده شد. ویروسها برخلاف باکتریها، قارچها و انگلها برای بقا مستقل نبوده و بهطور کامل به سلولهای زنده یوکاریوتی یا پروکاریوتی وابسته هستند و انگلهای درونسلولی اجباری میباشند. ویروسها برای همانندسازی وابسته به دستگاههای بیوشیمیایی سلول میزبان میباشند.
دانشمندان منشأ تکاملی ویروسها را دارای دو تئوری میدانند:
تئوری ۱– ذرات ویروسی ممکن است از ترکیبات اسید نوکلئیک DNA یا RNA سلول میزبان منشأ گرفته باشند.
تئوری ۲– ویروسها را حاصل از تغییر شکل ذرات داخلی سلول میدانند. ویروسها کوچکترین عوامل عفونی با قطر بین ۲۰ تا حدود ۳۰۰ نانومتر میباشند.
ویروسها فاقد ریبوزوم، میتوکندری و ارگانهای دیگر سلولی هستند و بهطور کامل به سلولهای زنده وابستهاند، لذا انگلهای درونسلولی اجباری بوده و توانایی تولید انرژی یا پروتئینها را بهصورت مستقل از سلول زنده ندارند. ژنوم ویروسها فقط از یک نوع اسید نوکلئیک تشکیل شده که میتواند RNA یا DNA باشد.
ویروسها قادرند در موجودات زنده مثل گیاه، جانور، حشره یا باکتری تکثیر یابند، لذا سه گروه بزرگ از ویروسها شامل ویروسهای حیوانی، گیاهی و باکتریایی وجود دارند. عوامل مؤثر در پیشرفت علم ویروسشناسی عبارتند از:
- ساخت میکروسکوپ الکترونی
- دسترسی به تکنیکهای خالصسازی ویروسها
- کشف پدیده هماگلوتیناسیون
- استفاده از تکنیکهای مولکولی
تعاریف مهم در ویروسشناسی
ویریون: ذره کامل ویروسی که حاوی ژنوم نوکلئیک اسید پوشیده شده در یک پوشش پروتئینی (کپسید) یا فاقد پوشش میباشد را ویریون میگویند.
کپسید: پوستهای از جنس پروتئین است که توسط ژنوم ویروس کد میشود و ژنوم ویروس را فرا میگیرد و مسئولیت محافظت از ژنوم را بعهده دارد. در بعضی از ویروسها در نفوذ به هسته سلول میزبان نقش دارد.
کپسومر: واحدهای تشکیلدهنده کپسید میباشند که بهعنوان واحدهای مرفولوژیک با میکروسکوپ الکترونی در سطح ذرات ویروسی با تقارن ۲۰ وجهی دیده میشوند و در دور ژنوم در ویروسهای با تقارن مارپیچی مشاهده میشوند.
پروتومر یا واحدهای ساختمانی: در ساختمان کپسید واحدهای پروتئینی تشکیلدهنده مشاهده میشوند که واحدهای ساختمانی نامیده میشوند. از کنار هم قرار گرفتن چند واحد ساختمانی که با نظم خاصی در کنار هم قرار میگیرند، ساختمان منظم کپسید شکل میگیرد.
پروتئین ماتریکس: بعضی از ویروسهای پوششدار دارای پروتئین واسط بین پوشش و نوکلئوکپسید خود میباشند که ماتریکس نامیده میشود. در ویروسهای پوششدار مثل هرپس ویروسها و آنفلوانزا ماتریکس وجود دارد.
تگومنت: در ویروس هرپس ساختار خاص ناقرینه و بیشکل بین کپسید و پوشش وجود دارد.
پپلومر یا spike (خار): در سطح پوشش ویروسها زائدههایی از جنس گلیکوپروتئین وجود دارد که پیلومر یا VAP (Virus attachment protein) یا (Ligand) نام داشته و واسطه بین اتصال ویروس و گیرنده ویروس در سطح سلول یا رسپتور میباشند. پیلومرها مهمترین آنتیژنهای ویروسی میباشند (شکل ۱).
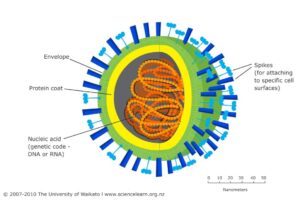
شکل ۱: بخشهای مختلف یک ویروس
شکلگیری کپسید:
در شکلگیری کپسید ویروس، پروتئینهای مجزا با پیوستن به هم، زیرواحدها (Subunits) را شکل میدهند و جمع زیرواحدها، واحدهای ساختمانی (Protomer) را ایجاد میکنند. از انسجام زیرواحدهای ساختمانی، کپسومرها ساخته میشوند و در نهایت پروکپسید و کپسید شکل میگیرد.
در بعضی از ویروسها شکلگیری کپسید همزمان با شکلگیری ژنوم ویروس اطراف آن انجام میشود، ولی در بعضی از ویروسها کپسید ابتدا به شکل نیمهکامل و خالی ساخته شده (پروکپسید) و بعداً ژنوم در داخل آن قرار میگیرد.
اجزای ویروس دارای پوشش
- – گلیکوپروتئینهای روی سطح پوشش ویروس
پروتئینهای سطحی ویروس (پیلومر) در دستگاه گلژی قنددار میشوند (گلیکوزیلاسیون) و تبدیل به گلیکوپروتئین میگردند و پپلومرها را تشکیل میدهند. قندهایی که به گلیکوپروتئینهای ویروس افزوده میشوند مربوط به سلول میزبان است و پروتئین آن را خود ویروسها تولید میکنند.
بعضی از این گلیکوپروتئینها توانایی اتصال به گلبولهای قرمز را دارا میباشند که به آنها هماگلوتینین (Hemagglutinin) میگویند. همچنین بعضی از این گلیکوپروتئینها میتوانند نقش فیوژن (پارامیکسوویروسها)، نورآمینیداز (آنفلوآنزا) و گیرنده FC (هرپس ویروسها) را داشته باشند.
ویروپورین: بعضی از ویروسها دارای پروتئینهای غشایی دیگری به نام ویروپورین هستند که پروتئینهای تشکیلدهنده کانالهای یونی بوده و در انتقال یونی نقش دارند.
VPU (Virion protein unique) در ویروس نقص ایمنی انسان (HIV)، p۷ در ویروس هپاتیت C (HCV)، ۶K در توگاویروسها، SH در پارامیکسوویروسها و M۲ در ویروس آنفلوانزا بهعنوان ویروپورین در ویروسهای دارای پوشش وجود دارند.
۲– پوشش (Envelope)
ویروسهای دارای پوشش، حاوی پوششهای لیپیدی میباشند که هنگام کامل شدن ویروس از غشاء سلولی به دست میآورند. ممکن است این پوشش از غشاء هسته (هرپس ویروس)، غشاء سیتوپلاسمی (پاراآنفلوآنزا) و یا از دستگاه گلژی (بونیاویروس) به دست آید.
۳– پروتئین ماتریکس:
در ویروسهای پوششدار مثل هرپسویروسها و آنفلوآنزا ماتریکس وجود دارد.
۴- کپسید:
با توجه به طبیعت اتصالهایی که در گردهمایی کپسومرها وجود دارد، اشکال مشخص از کپسیدها شکل میگیرد و از نظر ظاهری دو نوع تقارن وجود دارد؛ تقارن بیستوجهی (Icosahedral) و تقارن مارپیچی (Helical). بعضی از ویروسها در قاعده این دو تقارن قرار ندارند که به آنها ویروسهای نامتقارن، پیچیده یا کمپلکس نیز میگویند (شکل ۲).
الف- تقارن بیستوجهی
در این تقارن کپسومرها بهصورت مثلث متساویالاضلاع قرار میگیرند. این تقارن بهترین فرم پایداری کپسید ویروس از لحاظ هندسی است. این بیست وجهی دارای ۳۰ ضلع (لبه) و ۱۲ رأس است.
ب- تقارن مارپیچی (هلیکال)
زیرواحدهای کپسید یا کپسومرها در ارتباط و اتصال نزدیک به ژنوم ویروسی میباشند و به شکل مارپیچی دیده میشوند. به مجموعه کپسید و اسید نوکلئیک ویروس، نوکلئوکپسید میگویند که بایستی درون پوشش حاوی لیپید (انولوپ) قرار گیرند. در این تقارن همزمانی تولید اسید نوکلئیک و اتصال کپسومر وجود دارد، ویروکپسید وجود ندارد. این نوع تقارن در ویروسهای حاوی DNA دیده نمیشود و مخصوص RNA ویروسها است.
ج- تقارن پیچیده یا کمپلکس
بعضی ویروسها دارای تقارن مشخص نیستند، بلکه ساختمان پیچیدهتری دارند و مجموعهای از ساختمانهای مکعبی و مارپیچی هستند.
پاکسویروسها دارای ساختمانی آجری شکل بوده و دارای بخش مرکزی و اجســــــــــام جانبی (Lateral body) میباشند. باکتریوفاژها نیز دارای ساختمان مرکب میباشند.
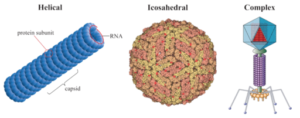
شکل ۲: اشکال مختلف از کپسیدها
۵- اسید نوکلئیک
ژنوم ویروس مرکزیترین قسمت ویروس است. ویروس دارای DNA یا RNA است (شکل ۳). DNA میتواند تکرشتهای یا دو رشتهای، خطی یا حلقوی باشد.
RNA میتواند سنس مثبت + شبیه (mRNA) و سنس منفی (-) و یا آمبیسنس (ambisense) (حاوی مناطق + و – ) باشد. ژنوم RNA ویروسها بهصورت هاپلوئید است، بهجز رتروویروسها که دیپلوئیدی هستند.
ژنوم سنس مثبت ویروس دارای خاصیتی همچون mRNA است و بعد از ورود میتواند در کنار ریبوزوم ترجمه شده و تولید پروتئین کند.
RNA این ویروسها عفونتزا محسوب میشود و این ویروسها همراه خود RNA پلیمراز ویروس را ندارند.
ژنوم این ویروسها در انتهای ‘۵ خود دارای Cap و در انتهای ‘۳ خود دارای دم پلی A (polyA) میباشند.
RNA ویروسهای دارای ژنوم پلاریته منفی فاقد خاصیت mRNA میباشند و بایستی ویروس درون خود آنزیم RNA پلیمراز ویروس را همراه داشته باشد تا ژنوم را به mRNA تبدیل کرده و مراحل تکثیر ویروس طی شود.
Ambisense خانواده آرناویروسها و بونیاویرسها دارای ژنوم مخلوط یا آمبیسنس هستند که نیمی از آن منفی و نیمی مثبت میباشد.
برخی از ویروسها از یک RNA تشکیل میشوند و برخی ژنوم بهصورت قطعهقطعه دارند مثل ویروس آنفلوانزا که دارای ژنوم متشکل از RNAهای ۷ تا ۸ قطعهای است.
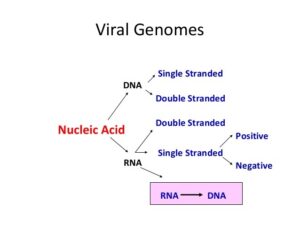
شکل ۳: انواع ژنوم در ویروسها
فراپژوهش